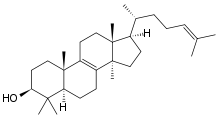
Cholesterol – organiczny
związek chemiczny
,
lipid
z grupy
steroidów
zaliczany także do
alkoholi
[5]. Jego pochodne występują w
błonie
każdej komórki zwierzęcej, działając na nią stabilizująco i decydując o wielu jej własnościach. Jest także prekursorem licznych ważnych steroidów takich jak
kwasy żółciowe
czy
hormony steroidowe
.
Potocznie cholesterolem nazywa się obecne w
osoczu krwi
pokrewne substancje lipidowe –
lipoproteiny
, w skład których między innymi wchodzi też cholesterol.
Stanowi on substrat do syntezy wielu ważnych biologicznie czynnych steroidowych cząsteczek:
W organizmie człowieka cholesterol występuje w tkankach i w osoczu krwi w postaci wolnej oraz w postaci zestryfikowanej długołańcuchowymi
kwasami tłuszczowymi
. Ogólna ilość cholesterolu w organizmie przeciętnego człowieka jest oceniana na 150–200 g, a wg innego źródła na jedynie 140 g, czyli 363 mmol[6].
Przestarzała nazwa cholesterolu to cholesteryna.
Budowa cząsteczki
Cholesterol zaliczamy do
steroidów
. Są to związki należące do
lipidów
(cechą łączące bardzo różne substancje tej grupy jest słaba rozpuszczalność w wodzie charakteryzująca też cholesterol umiejscawiający się w organizmie w lipoproteinach albo błonach biologicznych - w środowisku hydrofobowym).
Jak w przypadku innych steroidów, cząsteczka cholesterolu posiada 3 pierścienie sześciowęglowe (
cykloheksanowe
, jeden z nich zawiera też wiązanie podwójne) i czwarty pięciowęglowy (
cyklopentanowy
)[7]. Oznacza się je kolejnymi literami alfabetu łacińskiego odpowiednio A, B i C oraz D. Pierścienie sześciowęglowe, przedstawiane na rysunkach zazwyczaj w postaci
sześciokąta foremnego
, w rzeczywistości przyjmują konformację krzesłową. Niesie to za sobą pewne konsekwencje. Między innymi z 12 podstawników pierścienia cykloheksanowego 6 położonych jest ekwatorialnie (równikowo, w bok), 6 kolejnych aksjalnie (3 w górę, 3 w dół względem teoretycznej płaszczyzny pierścienia) i te właśnie cechuje duże stłoczenie przestrzenne. Wobec tego atomy węgla tworzące sąsiadujące pierścienie leżą zwykle aksjalnie, a także w położeniu trans względem siebie (jedna z grup skierowana jest lekko do góry, druga nieznacznie w stronę przeciwną, podobnie, jak w przypadku trans-dekaliny[7]). Obie grupy metylowe ułożone aksjalnie niejako wystają na sztorc nad płaszczyzną pierścieni (są w położeniu β)[8]. Grupa hydroksylowa zajmuje położenie ekwatorialne[8].
Ta skomplikowana cząsteczka posiada aż 8 asymetrycznych (chiralnych) atomów węgla zwanych centrami stereogenicznymi. Są to atomy o numerach 3, 8, 9, 10, 13, 14, 17, 20. Teoretycznie korzystając z wzoru, wg którego maksymalna liczba izomerów wynosi 2n, gdzie n - liczba atomów chiralnych, można wyliczyć, że związek taki powinien mieć 28=256 izomerów optycznych, wśród których wyróżnilibyśmy 128 par
enancjomerów
, substancje należące do różnych par byłyby zaś względem siebie
diastereoizomerami
. Jednakże, jak to zazwyczaj ma miejsce w przypadku organizmów żywych, występuje w nich tylko jeden. Poza tym należy pamiętać, że mamy do czynienia z cząsteczką o licznych skondensowanych pierścieniach i wchodzące w grę naprężenia steryczne uniemożliwiają istnienie wielu z teoretycznych izomerów[9].
Reakcje chemiczne
Cholesterol ulega tym reakcjom chemicznym, co grupy funkcyjne, które posiada. Możemy więc wnioskować, że będzie reagował jak
alkohole
(grupa hydroksylowa przy trzecim atomie węgla) lub jak
alkeny
(wiązanie podwójne między piątym i szóstym atomem węgla).
Addycja do wiązania podwójnego
Wiązanie podwójne cholesterolu, położone, jak już było wspominanie, pomiędzy C5 i C6, może uczestniczyć w reakcjach addycji. Dla przykładu cholesterol reaguje z
bromem
(Br2). Oba atomy chlorowca przyłączają się analogicznie, jak w przypadku prostych alkenów, nie zważając na resztę cząsteczki[10].
Podobnie też przyłączają się chlorowcowodory.
Utlenianie
Grupa hydroksylowa
cholesterolu, jak każda drugorzędowa grupa alkoholowa, utlenia się do
karbonylowej
dzięki zastosowaniu utleniaczy, jak np. odczynnik Jonesa. Powstały produkt zaliczyć można do
ketonów
, dlatego też nosi on nazwę 5-cholesten-3-on[11].
Biosynteza
Pula cholesterolu organizmu człowieka pochodzi ze źródeł pokarmowych (egzogennych), w zależności od diety 300–500 mg dziennie oraz z biosyntezy (cholesterol endogenny) 700–900 mg dziennie, a wg innych źródeł nawet 3 g[6]. Produkcja zachodzi we wszystkich komórkach jądrzastych, lecz głównie w
wątrobie
(60-70%),
jelitach
(15%) oraz w skórze (5%). Do zastosowań przemysłowych jest on pozyskiwany ze źródeł naturalnych, gdyż koszt jego syntezy jest za wysoki.
Endogenny cholesterol syntetyzowany jest z tego samego źródła, co kwasy tłuszczowe: z acetylo-CoA. Tworzenie tej cząsteczki z fragmentów dwuwęglowych przebiega w kilku etapach[12].
Utworzenie mewalonianu
Początkowa reakcja syntezy cholesterolu z acetylo-CoA nie różni się zbytnio od tej zachodzącej w przypadku wspomnianych już kwasów tłuszczowych. Dwie cząsteczki acetylo-CoA reagują ze sobą w reakcji katalizowanej przez
enzym
tiolazę cytozolową. W rezultacie powstaje acetoacetylo-CoA i wolny
koenzym A
. Acetoacetylo-CoA może też powstać w inny sposób[12].
Acetoacetylo-CoA kondensuje z kolejną cząsteczką acetylo-CoA. Tym razem funkcję katalizatora pełni syntaza HMG-CoA. Kolejny wolny koenzym A ulega odszczepieniu, głównym produktem reakcji jest zaś 3-hydroksy-3-metyloglutarylo-CoA (β-hydroksy-β-metyloglutarylo-CoA, w skrócie HMG-CoA)[12].
3-hydroksy-3-metyloglutarylo-CoA ulega redukcji, dzięki czemu odłącza się ostatnia, trzecia cząsteczka koenzymu A. Równoważników redukcyjnych (atomów
wodoru
) dostarcza
NADPH
, czyli zredukowana postać fosforanu dinukleotydu nikotynoamidoadeninowego. Utlenia się ona do NADP+. Proces katalizuje
reduktaza HMG-CoA
. W jego efekcie powstaje
mewalonian
[12].
Wspomnieć należy, że ten właśnie etap podlega skomplikowanej regulacji zarówno naturalnej, jak i przy użyciu środków farmaceutycznych.
Statyny
– grupa leków obniżających poziom cholesterolu – kształtem cząsteczki przypominają substrat i prowadzą kompetycyjną inhibicję enzymu[12].
Stworzenie fragmentu pięciowęglowego
Izopren
, od którego pochodzi jednostka pięciowęglowa
Mewalonian posiada 6 atomów węgla, powstał bowiem z 3 reszt acetylowych. Do syntezy pierścienia steroidowego używana jest jednaj grupa pięciowęglowa, tzw. jednostka
izoprenoidowa
nazwana tak od węglowodoru posiadającego podobny szkielet węglowy. Warto zaznaczyć, że z jednostek takich składa się wiele substancji zwanych ogólnie terpenami, jak np.
kamfora
, a jej polimerami są
kauczuk
czy
gutaperka
. Tak czy inaczej mewalonian musi pozbyć się jednego atomu węgla[12].
Wbrew pozorom jest to bardzo kosztowne energetycznie, a zachodzi dzięki trzem działającym po sobie
kinazom
. Jako pierwsza bierze w tym udział kinaza mewalonianowa, fosforylując substrat przy piątym atomie węgla – powstaje 5-fosforan mewalonianu, który zostaje ufosforylowany po raz wtóry przez kinazę fosfomewalonianową, która z kolei czyni z niego 5-difosforan mewalonianu. Ten zaś ulega trzeciemu, ostatniemu już przeniesieniu grupy ortofosforanowej, dzięki czemu utworzeniu ulega 3-fosfo-5-difosforan mewalonianu. Dzieje się to dzięki kinazie difosfomewalonianowej. Fosforylacje te zużyły trzy cząsteczki
ATP
, czyniąc z nich 3
ADP
[12].
Utworzony tym sposobem 3-fosfo-5-difosforan mewalonianu ulega
dekarboksylacji
, oprócz dwutlenku węgla odłączając także jedną z grup fosforanowych. W efekcie powstaje więc difosforan izopentenylu zwany także izopentenylodifosforanem. Przejście to przeprowadza dekarboksylaza difosfomewalonianowa[12].
Łączenie jednostek izoprenoidowych
Część cząsteczek difosforanu izopentenylu ulega izomeryzacji katalizowanej przez izomerazę izopentenylodifosforanową do difosforanu 3,3-dimetyloallilu. Cały ten proces polega po prostu na zmianie położenia wiązania podwójnego[12].
Difosforan izopentenylu i difosforan 3,3-dimetyloallilu kondensują dzięki cis-prenylotransferazie, tworząc difosforan geranylu. Związek ten posiada już 10 atomów C[12].
Difosforan geranylu łączy się z kolejną cząsteczką difosforanu izopentenylu, dzięki czemu powstaje difosforan farnezylu o 15 atomów C. Może on wejść w kilka różnych szlaków metabolicznych, np. tworząc dolichol albo łańcuch boczny
ubichinonu
. By stać się w końcu cholesterolem albo innym sterydem, musi on jednak zostać połączony z drugą taką samą cząsteczką przez syntetazę skwalenu. Potrzebny jest także NADPH. W rezultacie powstaje liczący 30 atomów węgla
skwalen
. Mechanizm tej reakcji obejmuje utworzenie difosforanu preskwalenu, który jest redukowany i odłącza pirofosforan[12].
Cyklizacja
Spośród wielu możliwych konformacji, jakie może przyjąć skwalen, niektóre szczególnie sprzyjają cyklizacji łańcucha. Wpierw jednak epoksydaza skwalenowa przy udziale
tlenu
, NADPH i
FAD
(dinukleotydu flawinoadeninowego)[12] utlenia pierwsze wiązanie podwójne licząc od początku łańcucha (terminalne[8]), tworząc pierścień trójczłonowy charakterystyczny dla
epoksydów
. Powstały epitlenek nazywamy oksydoskwalenem[12] albo tlenkiem cholesterolu[8].
Następnie do akcji wkracza lanosterolocyklaza oksydoskwalenowa (cyklaza oksydoskwalen: lanosterol). Dzięki niej następuje zamknięcie się łańcucha w trzy pierścienie cykloheksanowe i 1 cyklopentanowy z przemieszczeniem grup metylowych przy C14 i C8 na odpowiednio C13 i C14 (tak więc C14 traci jedną grupę metylową i zyskuje nową). W efekcie powstaje 30-węglowy związek zwany
lanosterolem
[12].
Mechanizm tego przekształcenia opiera się na
karbokationach
. Uprotonowanie atomu węgla mającego stać się czwartej w cząsteczce cholesterolu czyni go podatnym na atak nukleofilowy atomu tworzącego położone blisko wiązanie podwójne (późniejszy C5). Ładunek dodatni gromadzi się więc na węglu sąsiednim (później 10), który przyciąga elektrony kolejnego wiązania podwójnego. W rezultacie karbokation przenosi się na przyszły C8 i sytuacja się powtarza. Przeniesienie tych trzech wiązań owocuje utworzeniem trzech pierścieni cykloheksanowych. Następnie obdarzony ładunkiem dodatnim węgiel, któremu przypisany zostanie nr 13, reaguje z przedostatnim wiązaniem podwójnym, tworząc pierścień pięcioczłonowy. Ładunek z C20 zamienia się miejscami z atomem wodoru z C17, a następnie C13. Przegrupowanie w obrębie karbokationu przenosi tam grupę metylową (będzie to C19), jej poprzednie miejsce (przy C14) zajmuje kolejna grupa metylowa złączona wcześniej z C8. Po kolejnej zmianie umiejscowienia ładunku karbokation rozpada się, przekazując kation wodoru jakiejś zasadzie i tworząc wiązanie podwójne. W ten sposób utworzona zostaje cząsteczka lanosterolu. Opisane tutaj po kolei kojne cyklizacje prawdopodobnie zachodzą jednocześnie, podobnie jak przegrupowania karbokationu i utrata H+[8].
Obróbka lanosterolu
Lanosterol posiada już grupę hydroksylową przy C3, ale dysponuje także nadmiarowymi grupami metylowymi, a jego wiązanie podwójne jest w złym miejscu. Musi więc nastąpić utlenienie wspomnianych grup do dwutlenku węgla, a wiązanie podwójne powinno zmienić pozycję. Jako kolejne stadia tego procesu zaproponowano:
- 14-demetylolanosterol
- zymosterol
- Δ7,24-cholestadienol
- desmosterol
Ten zaś pod wpływem Δ7,24-reduktazy zamienia się w cholesterol[12].
Jednakże pamiętać należy, że taka możliwość stanowi jedynie jedną z kilku. Nie ustalono bowiem dotychczas jednoznacznie, w jakiej kolejności zachodzą po sobie poszczególne przemiany[12].
Wchłanianie
Cholesterol egzogenny pochodzi z diety, przy czym zazwyczaj dzienne spożycie oscyluje w granicach 0,5-1 g[13]. Związek ten występuje obficie w pokarmach zwierzęcych, natomiast śladowe wręcz ilości można znaleźć w pożywieniu roślinnym. Bardzo dużą jego ilość zawiera żółtko
jaja
[13].
Jednak nie cały wchłaniany w przewodzie pokarmowym cholesterol pochodzi z konsumpcji. Związek ten dostaje się do dwunastnicy razem z
żółcią
wydzielaną przez
wątrobę
(jego masę szacuje się na 2 g dziennie), a także ze złuszczonym nabłonkiem wyściełającym przewód trawienny (dobowo około 0,5 g). W rezultacie blisko połowa wchłanianego cholesterolu nie pochodzi z pożywienia[13].
Z uwagi na charakterystyczną dla lipidów niską
rozpuszczalność
w wodzie cholesterol przebywa w świetle jelit w micelach mieszanych. Do wchłaniania cholesterolu konieczna jest więc obecność soli kwasów żółciowych[13].
Niezależnie od egzogennego lub endogennego pochodzenia najważniejszy ze steroidów wchłaniany jest w większości w proksymalnej części
jelita cienkiego
[13]. Procesowi temu ulega w formie wolnej i dopiero później zostaje zestryfikowany kwasami tłuszczowymi[13] o długich łańcuchach węglowodorowych.
Krążenie
Wchłonięty w jelitach cholesterol zostaje rozdzielony na dwie pule. Większa, stanowiąca około 70%, jako składnik
chylomikronów
przepływa najpierw przez naczynia limfatyczne, by w końcu znaleźć się w żyłach. Mniejsza część trafia natomiast do
żyły wrotnej
[13].
W ten sposób cholesterol trafia do
wątroby
, skąd jego część zostanie ponownie wydzielona do dwunastnicy[13].
Żółć zawiera bowiem cholesterol w postaci zarówno wolnej, jak i estrów. Jego stężenie szacuje się na 1,6-4,4mnol/dm3, co w przeliczeniu na gramy daje nam od 0,6 do 1,7 g/dm3. Tworzą się tam mieszane micele zawierające oprócz cholesterolu także sole kwasów żółciowych i fosfolipidy, wśród których przeważa
fosfatydylocholina
. Prawidłowo kwasów żółciowych powinno być 20-30 razy więcej, niż cholesterolu. W sytuacji, gdy ten stosunek spada do 13, rośnie ryzyko strącania się mikrokryształków cholesterolowych, co owocuje tworzeniem złogów. Ich powstawanie odbija się niekorzystnie na zdrowiu[6].
Znaczenie biologiczne
Schemat steroidogenezy (syntezy hormonów sterydowych)
Cholesterol odgrywa kluczową rolę w wielu procesach
biochemicznych
, m.in.: syntezie witaminy D3 oraz hormonów o budowie sterydowej takich jak
kortyzon
,
progesteron
,
estrogeny
i
testosteron
. Jego obecność w błonach komórek nerwowych
mózgu
ma duże znaczenie dla funkcjonowania
synaps
. Istnieją też doniesienia, że odgrywa on dużą rolę w działaniu
systemu immunologicznego
(odpornościowego).
Wątroba produkuje ok. 1 g cholesterolu dziennie w żółci. Niektóre badania wykazały, że cholesterol może działać jako antyoksydant[14].
Żółć
zmagazynowana w
pęcherzyku żółciowym
i pomagająca trawić
tłuszcze
jest ważna dla absorbowania witamin rozpuszczalnych w tłuszczach (
A
,
D
,
E
i
K
). Jest głównym prekursorem witaminy D i
hormonów steroidowych
, które zawierają
kortyzol
i
aldosteron
(w gruczołach nadnerczowych),
progesteron
,
estrogeny
,
testosteron
i ich pochodne. Dostarcza on szkielet strukturalny do
biosyntezy
wszystkich
steroidów
. W
mielinie
okrywa on i izoluje nerwy, wspomagając przepływ impulsów nerwowych.
Cholesterol także bierze udział w formowaniu
raftów lipidowych
i membrany plazmowej. Także redukuje przepuszczalność błony komórkowej dla
kationów wodoru
i
sodu
[15].
Cholesterol jest istotny dla struktury i funkcjonowania wklęsłych
kaweol
i wgłębienia osłon
klatrynowych
, włączając endocytozyny kaweolozależne i klatrynozależne. Rola cholesterolu w takich endocytozynach może być badana za pomocą
cyklodekstryny
(MβCD) usuwającej cholesterol z błony komórkowej.
Cholesterol a choroby układu krążenia
Potocznie "cholesterolem" nazywa się pokrewne substancje lipidowe występujące w osoczu krwi –
lipoproteiny
, w skład których, między innymi wchodzą grupy cholesterolowe. Tzw. zły (niskiej gęstości –
LDL
) i dobry (wysokiej gęstości –
HDL
) cholesterol to w rzeczywistości lipoproteiny – czyli dość złożone
polipeptydy
zawierające cholesterol wolny i zestryfikowany. Są one formą transportową cholesterolu. Udowodniono, że zaburzenia gospodarki lipidowej, którego miarą jest wzrost stężenia cholesterolu w surowicy krwi, jest istotnym czynnikiem ryzyka wystąpienia
miażdżycy
i chorób układu sercowo-naczyniowego takich jak choroba wieńcowa i
zawał serca
,
udar mózgu
i miażdżyca zarostowa kończyn dolnych.
Nadmiar cholesterolu szkodzi nie tylko układowi krwionośnemu - związek ten stanowi główny składnik kamieni żółciowych[12].
Normy zawartości we krwi
Na podstawie badań epidemiologicznych ustalono, że podwyższony poziom cholesterolu w surowicy krwi, czyli
hipercholesterolemia
, jest jednym z podstawowych czynników ryzyka wystąpienia chorób układu sercowo-naczyniowego[16].
Oznaczanie stężenia cholesterolu z podziałem na frakcje pozwala w lepszy sposób ocenić negatywny wpływ hypercholesterolemii na zdrowie. Należy zaznaczyć, że hypercholesterolemia jest tylko jednym z czynników ryzyka i jego znaczenie należy oceniać w powiązaniu z innymi czynnikami ryzyka.
Ustalono, że stężenie frakcji lipoprotein o niskiej gęstości (LDL) we krwi koreluje dodatnio z ryzykiem wystąpienia choroby niedokrwiennej serca, natomiast dużej gęstości (HDL) ujemnie[16]. Z tego względu przyjęło się nazywanie lipoproteiny niskiej gęstości – "złym" cholesterolem, a lipoproteiny dużej gęstości – "dobrym" cholesterolem.
Związek między podwyższonym stężeniem cholesterolu a ryzykiem choroby niedokrwiennej serca ma charakter ciągły i półlogarytmiczny.
Do oceny ryzyka chorób układu sercowo-naczyniowego wykonuje się badanie nazywane lipidogramem, które obejmuje:
- stężenie cholesterolu całkowitego,
- stężenie cholesterolu HDL,
- stężenie cholesterolu LDL,
- stężenie trójglicerydów.
Wyliczany jest także stosunek stężenia cholesterolu całkowitego do stężenia HDL cholesterolu.
Normy dla stężenia cholesterolu całkowitego (TC - ang. Total Cholesterol)we krwi są następujące:
- Norma: < 200
m
g
/
dl
(< 5,2 m
mol
/
l
),
- Poziom podwyższony: 200–250 mg/dl (5,1-6,5 mmol/l),
- Poziom znacznie podwyższony: > 250 mg/dl (>6,5 mmol/l).
Normy dla stężenia "dobrego" i "złego" cholesterolu są następujące:
- HDL ("dobry" cholesterol) – wyższe wartości stanowią lepszy wynik,
- Norma u mężczyzn 35–70 mg/dl (0,9-1,8 mmol/l),
- Norma u kobiet 40–80 mg/dl (1,0-2,1 mmol/l),
- LDL ("zły" cholesterol) – niższe wartości stanowią lepszy wynik,
- Norma: < 135 mg/dl (< 3,5 mmol/l),
- Poziom podwyższony: 135–155 mg/dl (3,5-4,0 mmol/l),
- Poziom znacznie podwyższony: > 155 mg/dl (> 4,0 mmol/l).
Amerykański „Narodowy program edukacji cholesterolowej” Narodowego Instytutu Serca, Płuc i Krwi zawiera wytyczne obniżania
statynami
poziomu LDL:
- do poniżej 70 mg/dl u osób z chorobami serca i u osób umiarkowanie zagrożonych tymi chorobami,
- do poniżej 100 mg/dl u pozostałych osób [17].
Ciekłokrystaliczne pochodne cholesterolu
Niektóre pochodne cholesterolu, zwłaszcza lipidy cholesterolowe, wykazują własności
ciekłokrystaliczne
, generują
fazę nematyczną
(
faza cholesterolowa
) skręconą (N*), nazywaną też czasem fazą cholesterolową. Pochodne te są stosunkowo tanie w produkcji i dlatego występują w mieszankach stosowanych w
wyświetlaczach ciekłokrystalicznych
, oraz barwnikach zdolnych do zmiany barwy pod wpływem zmian temperatury.
Lipid cholesterylowy – obecny w błonach komórkowych i posiadający własności ciekłokrystaliczne
Przypisy
- ↑ Władysław Zygmunt Traczyk, Andrzej Trzebski, Andrzej Godlewski: Fizjologia człowieka z elementami fizjologii stosowanej i klinicznej. Warszawa: Wydawnictwo Lekarskie PZWL, 2007, s. 942. .
- ↑
Cholesterol – podsumowanie
(
ang.
). PubChem Public Chemical Database.
- ↑
Cholesterol – karta leku (EXPT00945)
(
ang.
). DrugBank.
- ↑
John McMurry
: Chemia organiczna. Wyd. 4. Warszawa: Wydawnictwo Naukowe PWN, 2000, ss. 300 tom = 1.
- ↑
John McMurry
: Chemia organiczna. Wyd. 4. Warszawa: Wydawnictwo Naukowe PWN, 2000, ss. 636 tom = 1.
- ↑ 6,0 6,1 6,2
Stanisław Konturek
: Czynności wątroby. W: Władysław Zygmunt Traczyk, Andrzej Trzebski, Andrzej Godlewski: Fizjologia człowieka z elementami fizjologii stosowanej i klinicznej. Warszawa: Wydawnictwo Lekarskie PZWL, 2007, s. 806. .
- ↑ 7,0 7,1
John McMurry
: Chemia organiczna. Wyd. 4. Warszawa: Wydawnictwo Naukowe PWN, 2000, ss. 138 tom = 1.
- ↑ 8,0 8,1 8,2 8,3 8,4
John McMurry
: Chemia organiczna. Wyd. 4. Warszawa: Wydawnictwo Naukowe PWN, 2000, ss. 1120-1124 tom = 2.
- ↑
John McMurry
: Chemia organiczna. Wyd. 4. Warszawa: Wydawnictwo Naukowe PWN, 2000, ss. 310 tom = 1.
- ↑
John McMurry
: Chemia organiczna. Wyd. 4. Warszawa: Wydawnictwo Naukowe PWN, 2000, ss. 71 tom = 1.
- ↑
John McMurry
: Chemia organiczna. Wyd. 4. Warszawa: Wydawnictwo Naukowe PWN, 2000, ss. 663 tom = 1.
- ↑ 12,00 12,01 12,02 12,03 12,04 12,05 12,06 12,07 12,08 12,09 12,10 12,11 12,12 12,13 12,14 12,15 12,16 Kathleem M. Botham, Peter A. Mayes: Synteza, transport i wydzielanie cholesterolu. W: Robert K Murray, Daryl K Granner, Victor William Rodwell, Franciszek Kokot, Zenon Aleksandrowicz: Biochemia Harpera ilustrowana. Warszawa: Wydawnictwo Lekarskie PZWL, 2008, ss. 282-285. .
- ↑ 13,0 13,1 13,2 13,3 13,4 13,5 13,6 13,7
Stanisław Konturek
: Układ trawienny. W: Władysław Zygmunt Traczyk, Andrzej Trzebski, Andrzej Godlewski: Fizjologia człowieka z elementami fizjologii stosowanej i klinicznej. Warszawa: Wydawnictwo Lekarskie PZWL, 2007, s. 798. .
- ↑ Smith LL. Another cholesterol hypothesis: cholesterol as antioxidant.. „Free radical biology & medicine”. 1 (11), ss. 47–61 (1991).
PMID 1937129
.
- ↑ Haines TH. Do sterols reduce proton and sodium leaks through lipid bilayers?. „Progress in lipid research”. 4 (40), ss. 299–324 (lipiec 2001).
PMID 11412894
.
- ↑ 16,0 16,1
Andrzej Szczeklik
(red.): Choroby wewnętrzne : podręcznik multimedialny oparty na zasadach EBM. T. 1. Kraków: Medycyna Praktyczna, 2005, ss. 123–4, 127–31. .
- ↑
Static over Statins: Should Young People without Cholesterol Problems Take Statins?
Scientific American
Linki zewnętrzne