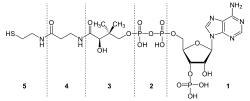
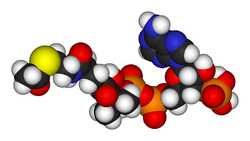
Struktura koenzymu A: 1: (3'-fosforyboza lub rybozo-3-fosforan i
adenina
) -
adenozyna
2: pirofosforan; 1+2: 3'-phosphoryliertes-Adenosindiphosphat 3: kwas pantoinowy, kwas dihydroksy-dimetylo-butanowy 4:
β-alanina
3+4:
kwas pantotenowy
5: β-merkaptoetyloamina, merkaptoetanoloamina, tioetanoloamina, cysteamina 3+4+5: Pantethein
Wzór strukturalny koenzymu A, wzór sumaryczny: C21H36N7O16P3S
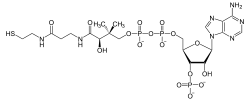
Wzór strukturalny Acetyl-CoA
Koenzym A (w skrócie CoA, czasem CoASH w celu uwidocznienia niepodstawionej
grupy tiolowej
) -
związek organiczny
powstający w
organizmie
z
adenozynotrifosforanu
,
pantotenianu
oraz
β-merkaptoetyloaminy
, służący jako przenośnik
grup acylowych
. Cząsteczkę koenzymu A związaną z resztą acylową nazywa się acylokoenzymem A (acylo-CoA). Najważniejszym z takich połączeń jest acetylokoenzym A (acetyl-CoA).
Acylo-CoA i Acetylo-CoA
Acylo-CoA czyli acylokoenzym A to połączenie koenzymu A z
resztą acylową
umożliwiające jej transport w organizmie. Acylo-CoA powstaje w wyniku
acylowania
grupy tiolowej
CoA:
- CoASH + RCOOH → CoAS~COR + H2O
Najważniejszym przykładem takiego połączenia jest acetylokoenzym A (Acetyl-CoA), tzw. aktywny octan - produkt
acetylowania
koenzymu A uczestniczący w wielu przemianach zachodzących w organizmie, np. w
cyklu kwasu cytrynowego
.
Acetylo-CoA odgrywa kluczową rolę w
metabolizmie
. Składa się z grupy octanowej (acylowej -COCH3)
związanej kowalencyjnie
z koenzymem A. Uczestniczy w przemianie tlenowej
sacharydów
w
Cyklu Krebsa
, w
syntezie kwasów tłuszczowych
oraz w syntezie
steroidów
.
Tworzenie acetylo-CoA w mitochondriach
Pirogronian
po wejściu do
mitochondriów
może ulec utlenieniu do CO2 i H2O (w
cyklu kwasów trójkarboksylowych
) bądź być wykorzystany do syntezy kwasów lub innych związków. Uprzednio jednak musi ulec utleniającej
dekarboksylacji
, tworząc "aktywny octan" - acetylo-CoA.
- pirogronian +
NAD
+ + CoA → acetylo-CoA +
NADH
+ H+ + CO2
- ΔG°= -33,5 kJ (-8,0 kcal/mol)
Faktyczny mechanizm reakcji jest znacznie bardziej złożony. W katalizującym kompleksie uczestniczą trzy
enzymy
główne (dehydrogenaza pirogronianowa, transacylaza liponianowa, dehydrogenaza liponianowa), pięć koenzymów (
pirofosforan tiaminy
,
kwas liponowy
, NAD+,
FAD
, CoA) oraz dwa enzymy regulujące (
kinaza
i fosfataza dehydrogenazy pirogronianowej).
Przebieg procesu zależy od stanu energetycznego komórki. Przy wysokim stężeniu
ATP
dehydrogenaza pirogronianowa przy udziale odpowiedniej kinazy przechodzi w nieaktywną formę ufosforylowaną, dzięki czemu cały proces ulega zahamowaniu. Przy niskim stężeniu
ATP
, o wysokim
ADP
oraz Ca2+ następuje pod wpływem swoistej fosfatazy defosforylacja dehydrogenazy pirogronianowej, z utworzeniem formy aktywnej enzymu.
Proces utleniającej dekarboksylacji
α-ketokwasów
jest źródłem czterech
wiązań wysokoenergetycznych
. Trzy tworzone są podczas utleniania poprzez łańcuch oddechowy powstałego NADH. Czwarte, powstałe na poziomie substratu zmagazynowane jest w postaci ~S-CoA. Przy dekarboksylacji pirogronianu wiązanie to nie jest źródłem ATP, jest jednak wykorzystywane w procesach syntezy. Proces utleniającej dekarboksylacji pirogronianu jest nieodwracalny.
Bibliografia
- Utlenianie kwasów tłuszczowych: ketogeneza. W: Harold A. Harper, Victor W. Rodwell, Peter A. Mayes Biochemia Harpera wyd. PZWL