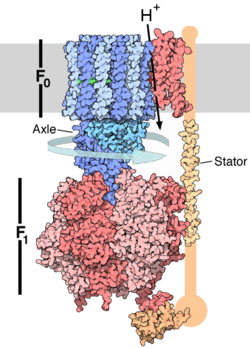
Schemat przedstawiający budowę syntazy ATP. Część F
O syntazy ATP, przez którą przepływa strumień protonów, zlokalizowana jest w wewnętrznej błonie mitochondrium. Część F
1, syntezująca ATP znajduje się na zewnątrz błony. Protony (H
+) przepływające kanałem utworzonym przez białka części F
O powodują, iż F
O obraca się. Podjednostka jest połączona z częścią F
O – a więc obraca się razem z nią. Tymczasem podjednostki, będące składnikami F
1 są unieruchomione, przez "zakotwiczoną" w błonie podjednostkę. Tak więc podjednostka obraca się (rotuje) wewnątrz cylindra powstałego z trzech podjednostek i trzech. Ponieważ jednak jest białkiem asymetrycznym, jego obroty "zmuszają" kolejne podjednostki do zmiany struktury –
konformacji
. Skutkiem zmian jest zróżnicowana, zmieniająca się w czasie, zdolność podjednostek do wiązania ATP, ADP
.
Syntaza ATP, syntetaza ATP – (
EC 3.6.3.14
)
enzym
katalizujący
reakcję wytwarzania związku wysokoenergetycznego –
ATP
z
ADP
i
fosforanu nieorganicznego
Pi. Energia niezbędna do syntezy pochodzi z
gradientu elektrochemicznego
i przekształcana jest w energię wiązań chemicznych podczas transportu
protonów
przez syntazę ATP.
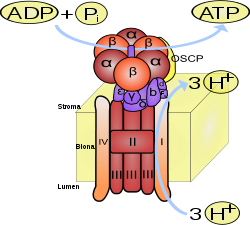
Enzym katalizuje reakcję:
ADP + Pi → ATP
Syntaza ATP znajduje się w wewnętrznej błonie
mitochondriów
, w błonach
tylakoidów
wewnątrz
chloroplastów
oraz w błonach komórkowych organizmów prokariotycznych. Enzym bierze udział w kluczowych procesach uzyskiwania energii przez organizmy żywe
fosforylacji oksydacyjnej
będącej głównym źródłem ATP wytwarzanego podczas
oddychania komórkowego
oraz
fosforylacji fotosyntetycznej
będącej źródłem ATP powstającego w fazie jasnej
fotosyntezy
.
Inhibitorem
syntazy ATP wykorzystywanym w badaniach biologicznych jest antybiotyk –
oligomycyna
.
Rola fizjologiczna
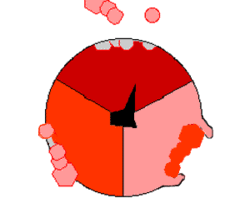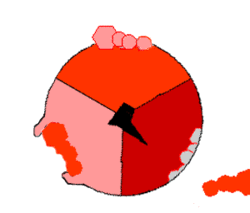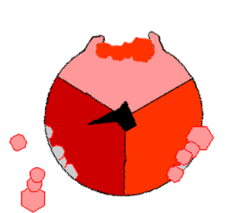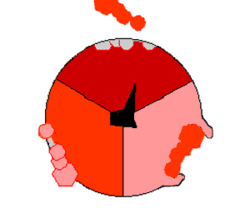
Mechanizm syntezy ATP. ATP – kolor czerwony, ADP i ortofosforan – kolor różowy, obracająca się podjednostka γ – kolor czarny.
Wewnątrz błony
mitochondrialnej
(u
eukariontów
) lub w
błonie komórkowej
(u
bakterii
) następuje utlenianie
NADH
do NAD+. Uwolnione
elektrony
uczestniczą w
łańcuchu oddechowym
, aby napędzić przeniesienie protonów w poprzek błony przez składające się na ten łańcuch odpowiednie przenośniki (pompy). W chloroplastach i błonach fotosyntetyzujących organizmów prokariotycznych protony przenoszone są dzięki energii uzyskanej poprzez pochłonięcie kwantów światła przez odpowiednie kompleksy.
ATP wytwarzane jest z
ADP
i
Pi
(reszty ortofosforanowej) w wyniku działania syntazy ATP. Rotacja jej odpowiedniego segmentu umożliwia syntezę ATP. Energia niezbędna do syntezy dostarczana jest przez
gradient elektrochemiczny
.
Budowa enzymu
Ze względu na miejsce występowania wyróżnia się trzy podstawowe kompleksy syntazy ATP.
Są to:
- Syntaza ATP mitochondrialna określana jako FoF1 lub MFoF1
- Syntaza ATP chloroplastowa określana jako CFoF1
- Syntaza ATP prokariotyczna
Wszystkie trzy syntazy ATP zaliczane są do grupy F-ATPaz i moją zbliżona budowę. Każda z nich składa się z dwóch domen. Pierwsza z nich – Fo – jest białkiem wewnątrzbłonowym tworzącym kanał jonowy dla jonów H+. Druga domena jest właściwą syntazą składająca się z kilkunastu polipeptydów tworzących kulista strukturę o średnicy ok. 100 Å.
Ewolucja
Uważa się, że ewolucja syntazy ATP zachodziła modułowo. Obie domeny wraz ze swoimi wartościowościami połączyły się zyskując nową funkcję. Domena F1 wykazuje znaczne podobieństwo do heksamerycznej
helikazy DNA
, a domena Fo jest podobna do kompleksów tworzących
motor molekularny
napędzający
wici
komórek.
Heksamer α3β3 tworzący domenę F1 podobnie jak helikaza DNA składa się z obracającego się pierścienia z otworem po środku. U obu enzymów obroty pierścienia umożliwiają pełnienie funkcji. Helikaza DNA porusza się po helisie DNA przy jednoczesnej hydrolizie nukleotydów. Domena F1 wykorzystuje zmiany konformacyjne podczas obrotu podjednostki γ do przeprowadzenia reakcji enzymatycznej.
Jony H+ przepływające przez domenę Fo w bardzo podobny sposób jak przy napędzaniu motoru molekularnego poruszającego wicią. Ich wspólną cechą jest pierścień składający się z wielu alfa-helikalnych białek, które obracają się w stosunku do innych białek zużywając jednocześnie gradient protonowy jako źródło energii. Jest to jednakże dość wątły związek, ogólna struktura motorów molekularnych jest zdecydowanie większa, zawierają 30 polipeptydów, w porównaniu do 10, 11, lub 14 znanych polipeptydów domeny Fo.
Modułowa teoria pochodzenia syntazy ATP sugeruje, że dwie domeny o niezależnych funkcjach, helikaza DNA posiadająca właściwości ATPazy i motor molekularny napędzany siłą protonomotoryczną, mogły się połączyć i doprowadzić do odwrócenia właściwości ATPazy powstałej z helikazy DNA. Dalszy rozwój prowadziłby do powstania kompleksy syntazy ATP znanej dzisiaj. Alternatywnie kompleks helikazy DNA i motoru molekularnego mógł wykazywać początkowo aktywność ATPazy, która przenosiła jony H+ zużywając ATP. Dalsza ewolucja kompleksy mogłaby doprowadzić do odwrócenia przeprowadzanej reakcji i powstania funkcji spełnianej przez syntazę ATP.
Numer EC 3.6.3.14
Bibliografia
- Boyer PD. The binding change mechanism for ATP synthase--some probabilities and possibilities.. „Biochimica et biophysica acta”. 3 (1140), ss. 215–50 (styczeń 1993).
PMID 8417777
.
- Boyer PD. The ATP synthase--a splendid molecular machine.. „Annual review of biochemistry”, ss. 717–49 (1997).
doi:10.1146/annurev.biochem.66.1.717
.
PMID 9242922
.
- Devenish RJ., Prescott M., Roucou X., Nagley P. Insights into ATP synthase assembly and function through the molecular genetic manipulation of subunits of the yeast mitochondrial enzyme complex.. „Biochimica et biophysica acta”. 2-3 (1458), ss. 428–42 (maj 2000).
PMID 10838056
.
- Velours J., Paumard P., Soubannier V., Spannagel C., Vaillier J., Arselin G., Graves PV. Organisation of the yeast ATP synthase F(0):a study based on cysteine mutants, thiol modification and cross-linking reagents.. „Biochimica et biophysica acta”. 2-3 (1458), ss. 443–56 (maj 2000).
PMID 10838057
.
- Gresser MJ., Myers JA., Boyer PD. Catalytic site cooperativity of beef heart mitochondrial F1 adenosine triphosphatase. Correlations of initial velocity, bound intermediate, and oxygen exchange measurements with an alternating three-site model.. „The Journal of biological chemistry”. 20 (257), ss. 12030–8 (październik 1982).
PMID 6214554
.
Zobacz też
Linki zewnętrzne